by Ben Best
Losing weight not only makes a person look good, it can reduce the danger of
getting heart disease, diabetes and
cancer [*1]. But
the health hazards and benefits of fats, carbohydrates and proteins — and their
effectiveness for diets and dieting — depend greatly on the type
of fat, carbohydrate and protein.
(See Macronutrients, Dieting and Health)
Dietary fat by itself, not just the body fat it produces, can be a health hazard. A study has shown that reducing dietary fat from 36% of total calories to 26% of total calories can significantly lower blood pressure within 8 weeks [*2]. Saturated fat in the diet can increase the risk of heart disease from atherosclerosis (fatty plaques on blood vessel walls) by raising blood cholesterol. Unsaturated fat is more likely to form free radicals by lipid peroxidation — which can lead to cancer and may accelerate aging. Therefore, both saturated and unsaturated fat can have health hazards. But every cell membrane in the body contains fat, and some of those fats cannot by synthesized — making it essential to obtain these fats from diet.
Some nutritionists have recommended substituting mono-unsaturated and poly-unsaturated fats for saturated fats, but another recommendation is to substitute protein and carbohydrate calories for fat calories [*3]. Fats (especially animal fats) are the primary vehicle by which pesticides enter the body. Some people might conclude that it would be a good idea to eliminate all fat from the diet. But eliminating all fat is not a good idea.
Your body needs fats to function properly. Nearly half of the dry weight of the brain is fat, and a quarter of this is cholesterol. Cholesterol is an essential part of sex hormones, bile acids, D vitamins and steroid hormones from the cortex of the adrenal gland — among other important substances. Cholesterol does not need to be eaten, however, because the liver and other tissues can manufacture cholesterol from saturated fats. But too many saturated fats result in excessively high blood levels of cholesterol that can end up being deposited in atherosclerotic plaques on blood vessels, leading to cardiovascular disease. High blood cholesterol also depresses the immune system and thereby increases the incidence of cancer [*4]. Excessive blood cholesterol is more often caused by eating too many saturated fats than by eating cholesterol itself.
Studies in the United States and Northern Europe have established that the incidence of coronary heart disease mortality is nearly two-and-a-half times higher for people with the highest 25% of blood cholesterol compared with people with the lowest 25%. Yet the coronary heart disease mortality for the same cholesterol levels is only one-third as great in Japan or the Mediterranean [*6]. A person in west Scotland having the same blood cholesterol levels as a person in Catalonia, Spain is 8-times more likely to die of coronary heart disease. So although cholesterol can be a factor in coronary mortality, it is not the only factor.
The term blood cholesterol is actually a reference to HDL (High-Density Lipoprotein), LDL (Low-Density Lipoprotein) and VLDL (Very Low-Density Lipoprotein). "Lipo" means "lipid", a general term that refers to all biological fats and oils. HDL has been called "good cholesterol" because it can pick up excess fats and carry them back to the liver. LDL (and especially VLDL) has been called "bad cholesterol" because it can become so overloaded with fats that the fats are dropped on blood vessel walls rather than carried to the cells where they are needed. LDL could also be called "good", because LDL supplies cells with fats needed for structure and function. In fact, LDL is the main carrier of oil-soluble substances (like Vitamin E) to body cells [*5]. LDL-cholesterol is just HDL-cholesterol with a larger load of surrounding fat. Exercise creates HDL from LDL by removing fat from LDL for use as energy. HDL has a multitude of cardioprotective actions in addition to its role in removing cholesterol from cells [CARDIOVASCULAR RESEARCH; Kontush, A; 103(3):341-349 (2014)].
Oxidation of LDL is what causes cholesterol to be deposited in plaque on blood vessel walls [*5]. The vulnerability of LDL cholesterol to oxidation depends on both the quantity of antioxidants in the blood and on the type of fatty acid in the LDL. Fruits, vegetables and supplements (especially Vitamin E with Vitamin C) will reduce LDL oxidation. The dietary fat that is most vulnerable to oxidation is linoleic acid [*7]. High blood levels of homocysteine thiolactone cause LDL to aggregate. Oxidized, aggregated LDL is readily attacked by macrophages to form atherosclerotic plaques on blood vessel walls [*8]. Small particle LDL cholesterol most readily causes atherosclerosis [JOURNAL OF ATHEROSCLEROSIS AND THROMBOSIS; Koha,S; 15(5):250-260 (2008)] because small particles are more easily oxidized and more easily cross blood vessel walls. So LDL particle size is a more critical indicator of atherosclerotic risk than total cholesterol or LDL cholesterol.
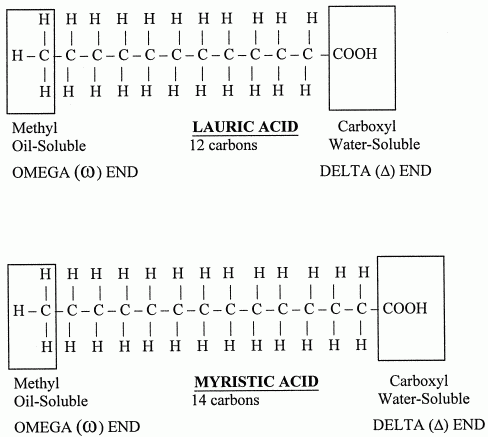
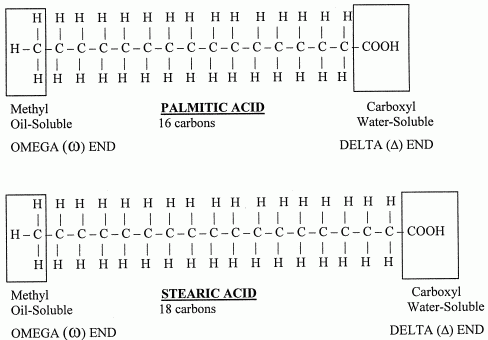
Aside from cholesterol, most other fat in the body is constructed from what is known as fatty acids. A fatty acid is a chain of carbon atoms (studded with hydrogen atoms) that has an acid group (carboxylic acid) at one end (the water-soluble end). The rest of the fatty acid is oil-soluble, with a methyl group at the other end. Fatty acids in the body usually exist unattached to any other molecule (free fatty acids), attached to glycerol in groups of three (triglycerides), or attached to phosphatidic acid molecules (phospholipids).
Fatty acids differ from one another not only by the number of carbons in their chain, but by the number of double-bonds between the carbon atoms. Fatty acids with only single-bonds are called saturated because they are "saturated" with as many hydrogen atoms as they can carry. Fatty acids with only one double-bond are called mono-unsaturated. And fatty acids with more than one double-bond are called poly-unsaturated.
Polyunsaturated fats have often been recommended to reduce coronary heart disease [*9]. But all saturated fats do not have the same effect on cholesterol synthesis in the liver. Only the saturated fats of chain-length 12, 14 and 16 (lauric acid, myristic acid and palmitic acid) have been shown to elevate blood cholesterol. Of these, myristic acid (high in coconut and palm oil) elevates cholesterol the most [*10]. Stearic acid (18-carbon, saturated) has been shown to lower cholesterol by 21% — even more than oleic acid (18-carbon, mono-unsaturated) which lowers LDL by 15% [*11].
Polyunsaturated fatty acids can be a health hazard because carbon-carbon double bonds can lead to free-radical formation and reactions with oxygen to form unstable lipid peroxide compounds containing the same unstable oxygen-oxygen bond found in hydrogen peroxide. Lipid peroxidation and free radicals can cause cancer and may accelerate aging. High rates of lung cancer among women in China have been associated with lipid peroxidized oils in fumes from cooking polyunsaturated vegetable oils in a wok [*12]. Hot oil in open air is subject to much lipid peroxidation. Fast-food restaurants that fry foods in the same oil all day serve lots of lipid peroxides to their customers.
Polyunsaturated "cis" fatty acids can be beneficial in cell membranes by preventing the tight packing of fatty acids in membranes — thereby making the membranes more "fluid". Membrane fluidity is important for optimal function of most cells in the body. But membrane fluidity is especially important on portions of cells that act as receptors for hormones or neurotransmitters. The typical North American eats three times as much saturated fat as unsaturated fat, yet animal experiments show that insulin receptor responsiveness is substantially improved when dietary unsaturated fat is greater than saturated fat [*13]. With aging, however, cell membrane fluidity declines in part because of increasing amount of cholesterol in the membranes, but more importantly because of free-radical oxidation [*14]. Antioxidants that protect cell membranes, like Vitamin E, are extremely valuable in opposing membrane oxidation.
| CIS ISOMER | TRANS ISOMER |
|---|---|
![[ <B><I>CIS</I> ISOMER</I></B> ]](Cis-2-butene.jpg)
|
![[<B><I>TRANS</I> ISOMER</I></B> ]](Trans-2-butene.jpg)
|
Fatty acid double-bonds come in two configurations known as cis (carbon chains on the same side of a double-bond) and trans (carbon chains on the opposite side of a double-bond). Most of the double-bonds made by biological systems have the cis configuration. It is the cis configuration of unsaturated fatty acids that prevents tight packing of fatty acids in membranes, and hence increases membrane fluidity.
Saturated fats (like butter or lard) and fatty acids with trans double-bonds (like margarine) tend to be solids at room temperature, whereas natural fatty acids with cis double-bonds (like vegetable oils) tend to be liquids. By artificially hydrogenating vegetable oils, the food processing industry reduces the number of double bonds and causes the formation of trans fatty acids. Hydrogenation results in margarines that are more solid and less vulnerable to rancidity. Hydrogenation results in peanut butter with a trans-fat-containing oil that does not separate from the peanut paste. But when trans fatty acids are incorporated into cell membranes, the membrane fluidity is reduced and the cells do not function as well. Not all trans fatty acids in the diet are due to food processing. For example, natural butter is 5% trans fat.
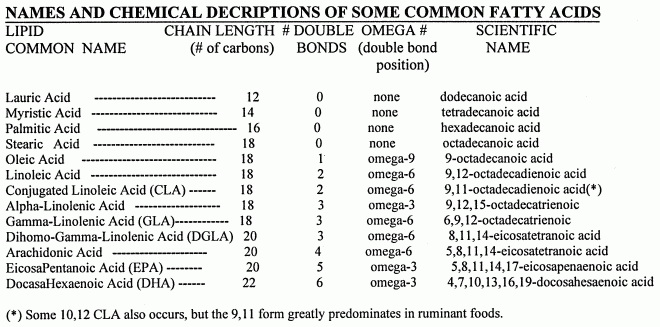
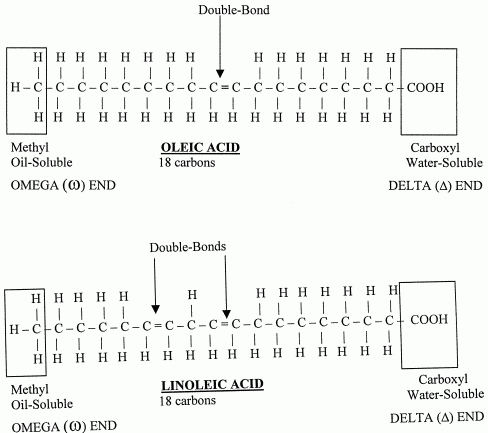
The human body can manufacture most of the fats it needs, including cholesterol, saturated fatty acids and unsaturated fatty acids. But there are fatty acids which cannot be manufactured in the body, and which must be obtained from dietary sources, for example linoleic acid and alpha-linolenic acid. These are essential fatty acids. Linoleic acid is an 18-carbon chain with 2 double-bonds, whereas alpha-linolenic acid is an 18-carbon chain with 3 double-bonds. The position of double-bonds in a fatty acid is critical to function, and this is especially true of double-bonds close to the methyl end. For long-chain fatty acids, the body's enzymes cannot add double-bonds near the methyl end.
A fatty acid has a carboxylic acid at one end and a methyl group at the other end. Carbon atoms in a fatty acid are identified by Greek letter on the basis of their distance from the carboxylic acid. The carbon atom closest to the to the carboxylic acid is the alpha (α) carbon, the next adjacent carbon is the beta (β) carbon, etc. In a long-chain fatty acid the carbon atom in the methyl group is called the omega (ω) carbon because omega is the last letter of the Greek alphabet. Because the closest double-bond to the methyl group in linoleic acid is 6 carbon atoms away from the methyl, linoleic acid is called an omega−6 (ω−6 or n−6) fatty acid. For alpha-linolenic acid, the double-bond closest to the methyl group is only 3 carbons away, so it is an omega−3 (ω−3 or n−3) ) fatty acid. The carbon next to the carboxylic acid is called the alpha carbon because alpha is the first letter of the Greek alphabet. But to confuse matters, the acid end of a fatty acid is called the delta (δ) end.
The configuration (geometry) at the double bond can be either
cis (adjacent hydrogen atoms on the same side of
the molecule) or trans (adjacent atoms on the opposite side).
Unsaturated fatty acids in the trans configuration are more
linear, more rigid and have a higher melting-point. The distinctiveness
of cis & trans fatty acids merit
unique names: cis-9-octadecanoic acid is oleic acid,
whereas trans-9-octadecanoic acid is elaidic acid.
(For more on trans fatty acids, see my essay
Are Trans-Fatty Acids a Health Hazard?.)
The body cannot make an omega−3 or omega−6 fatty acid because human metabolism cannot add a double-bond to a fatty acid that is more than 9 carbons away from the delta end. For the same reason, the body cannot convert an omega−3 to an omega−6 fatty acid, or vice-versa. But the body can make omega−9 fatty acids. And the body can add more double-bonds closer to the delta end of omega−3 and omega−6 fatty acids.
Two distinct families of essential fatty acids exist in the human body: the omega−3 family and omega−6 family. The omega−3 family (including EPA and DHA) comes from alpha-linolenic acid, and the omega−6 family (including arachidonic acid) comes from linoleic acid. Each family is the result of increasing chain length and of forming double-bonds from one of these two essential fatty acids. The two families compete for the same enzymes for forming double bonds (desaturase enzymes) and enzymes for lengthening the carbon chain (elongase enzymes). Elongase enzymes always add carbon atoms (in pairs) to the delta end of the fatty acid.
Whereas unsaturated fatty acids are denoted by the position of the
first double-bond from the methyl end (omega,ω) of the
fatty acid chain, the desaturase enzymes are denoted by the position
from the carboyl end (delta,Δ) at which the double-bond
is added. In synthesizing DHA (22:6ω−3) from
EPA (20:5ω−3) an elongase produces a 24-carbon
fatty acid chain, which is desaturated by Δ−6−desaturase
and then shortened to the 22-carbon DHA, which leaves the saturation
in the Δ−4 position [LIPIDS; Sprecher,H; 34:S153-S156
(1999)]. Less than 5% of dietary alpha-linolenic acid is converted
to DHA [CURRENT OPINION IN CLINICAL NUTRITION AND METABOLIC CARE;
Brenna,JT; 5(2):127-132 (2002)].
![[The Production of EFA Derivatives ]](efaprod.jpg)
|
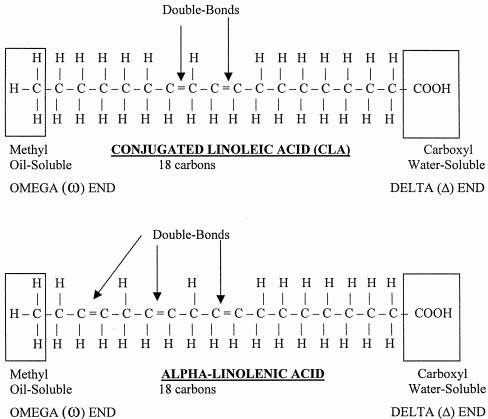
Forming a double-bond at the 9th carbon of linoleic acid results
in gamma-linolenic acid (GLA). Like alpha-linolenic acid, gamma-linolenic
acid has 3 double-bonds. But gamma-linolenic acid is an omega−6, whereas
alpha-linolenic is an omega−3 fatty acid. The words "alpha" and "gamma"
in this case
have no chemical meaning. The confusing terms "alpha-linolenic" and
"gamma-linolenic" are common names, not scientific names, and
are meaningless in the context of current scientific naming conventions.
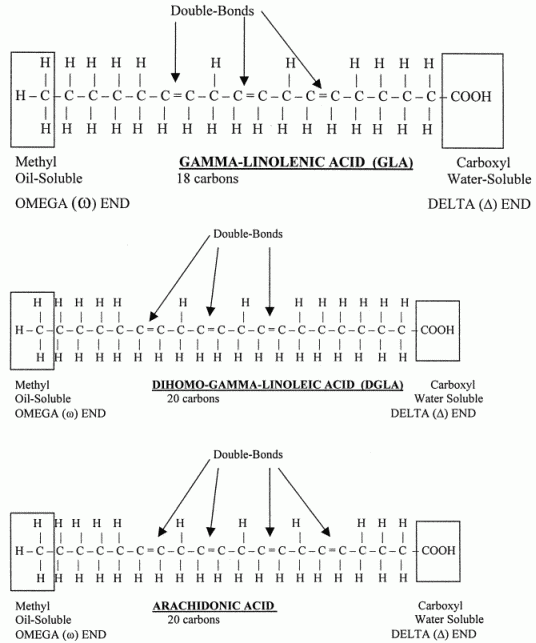
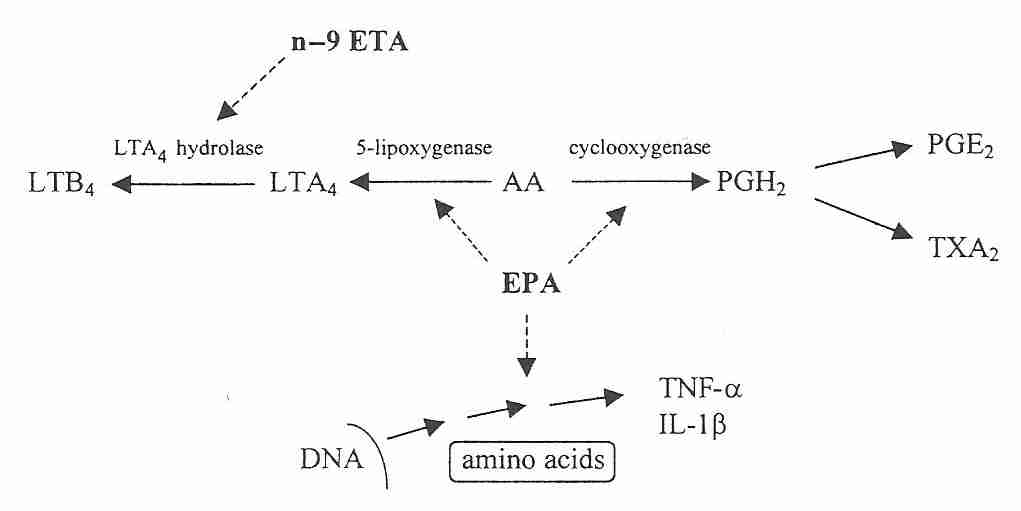
Another important product of linoleic acid is arachidonic acid. Arachidonic acid is a 20-carbon omega−6 fatty acid with 4 double-bonds. Arachidonic acid, in turn, gives rise to a whole group of 20-carbon, biologically-important substances known as the eicosanoids (eicosa- is Greek for "20"), including prostaglandins, thromboxanes, lipoxins and leukotrienes — which affect immunity, inflammation and blood clotting (among other actions). But omega−3 fatty acids can also produce eicosanoids.
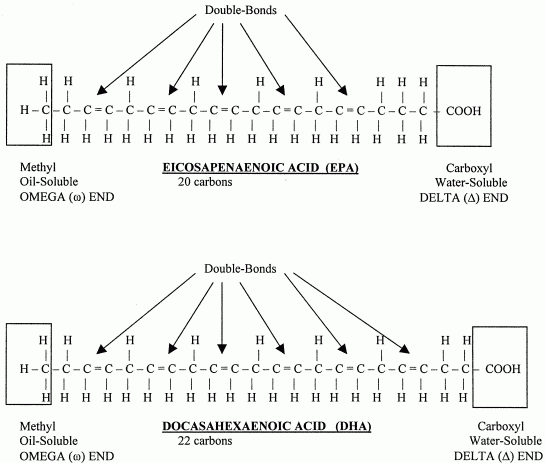
Noteworthy members of the omega−3 family of fatty acids manufactured from alpha-linolenic acid are EicosaPentaenoic Acid (EPA) and DocasaHexaenoic Acid (DHA). A pentaenoic acid has 5 double-bonds. A hexaenoic acid has 6 double-bonds. EPA is a 20-carbon chain fatty acid, whereas DHA is a 22-carbon chain fatty acid. Like arachidonic acid, EPA gives rise to its own class of eicosanoids. The EPA-generated eicosanoids are in the omega−3 family, as distinct from the omega−6 eicosanoids derived from arachidonic acid.
The primary source of omega−6 fatty acids in the diet is linoleic acid from the oils of seeds and grains. Sunflower, safflower and corn oil are particularly rich sources of linoleic acid, which is at the root of the omega−6 fatty-acid family. Evening primrose oil and borage oil are high not only in linoleic acid, but the omega−6 derivative gamma-linolenic acid (GLA). Avocado is 15-20% oil — mainly monosaturated, but also high in linoleic acid. (Avocado has the highest fat content and the highest fiber content — soluble as well as insoluble — of any fruit.)
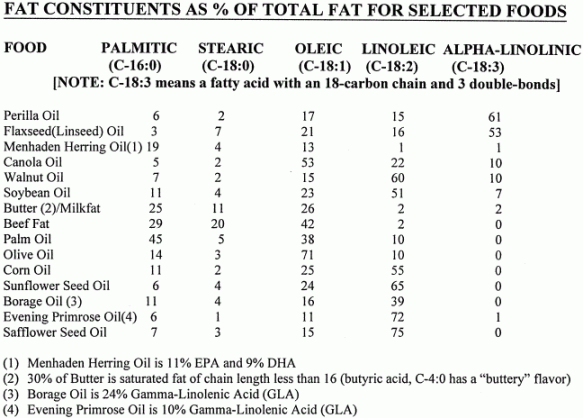
Alpha-linolenic acid, the primary dietary source of omega−3 fatty acids in the diet, is frequently found in green leaves. The leaves and seeds of the perilla plant (widely eaten in Japan, Korea and India) are the richest plant source of alpha-linolenic acid, although linseed oil is also a rich source. Fish oil contains very little alpha-linolenic acid, but is rich in the omega−3 derivatives EPA and DHA. Fish are at the top of a food chain based on phytoplankton (algae) that manufacture large amounts of EPA & DHA. Salmon and herring are about 4 or 5 times richer in EPA & DHA than cod [BIOCHEMICAL PHARMACOLOGY; Russo,GL; 77(6):937-946 (2009)]. Fish oils have been shown to improve insulin sensitivity and reduce inflammation in animal studies. In humans, higher intake of omega-3 fatty acids results in fewer of the pro-atherogenic small LDL particles [INTERNATIONAL JOURANAL OF MOLECULAR SCIENCES; 25:2104 (2022)]. DHA is superior to EPA in decreasing blood pressure, heart rate, and platelet aggregation [PROCEEDINGS OF THE NUTRITIONAL SOCIETY; Cottin,SC; 70(2):215-231 (2011)]. Fish can be high in toxic methylmercury, however. (For more information, see my essay Is Mercury in Fish a Health Hazard?) Possibly this is the reason fish oil supplements have not been shown to reduce cardiovascular disease in humans [INTERNATIONAL JOURANAL OF MOLECULAR SCIENCES; 19:3103 (2018)].
It has been estimated that thousands of years ago the diet of human hunter-gatherers consisted of approximately equal parts of omega−3 and omega−6 essential fatty acids [*15]. Since the beginning of agriculture ten thousand years ago there has been a steady increase in omega−6 at the expense of omega−3 fat in the human diet. This process accelerated about 50 years ago as cattle began to be fed increasingly on grains rather than grass. Recommendations by nutritionists to eat margarine rather than butter (polyunsaturated rather than saturated fats) increased the trend toward pro-inflammatory omega−6 and trans fat consumption. Currently, the ratio of omega−6 to omega−3 fatty acids in the American diet is 7−to−1 or more. There have been reasons to believe that this imbalanced essential fatty acid ratio has led to increased cancer, heart disease, allergies, diabetes and other afflictions. Much of the reason for this lies in the membranes of our cells. Nonetheless, these reasons are now being challenged: [JOURNAL OF PREVENTIVE CARDIOLOGY; Khandelwal,S; 2:325 (2013) and JOURNAL OF THE ACADEMY OF NUTRITION AND DIETETICS; Johnson,GH; 112:1029 (2012)].
The saturated fatty acid pentadecanoic acid (C15:0) has emerged as a previously unrecognized essential fatty acid. To evaluate this proposition it has been compared to eicosapentaenoic acid (EPA) [PLOS ONE; Venn-Watson S; 17(5):e0268778 (2022)] and shown to be superior to EPA. Pentadecanoic acid is present in 1% to 3% of dairy fat [NUTRIENTS; Venn-Watson S; 15:4607 (2022)], and compares favorably with compounds intended to extend lifespan and healthspan. A metabolite of pentadecanoic acid shows it to be an endocannaboid [SCIENTIFIC REPORTS; Venn-Watson S; 12:13717 (2022)].
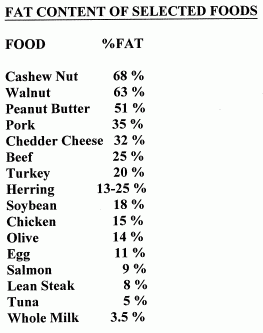
The table below classifies the oils found in common dietary foods on the basis of fat content, notably omega−3 (anti-inflammatory), omega−6 (pro-inflammatory), palmitate (the most harmful saturated fat) and omega−9. The table is roughly orders in descending order of healthfulness based on the above criteria (anti-inflammatory omega−3 is preferable to pro-inflammatory omega−6, and palmitate is undesirable). Palmitic acid (palmitate) induces insulin resistance in brain as well as in muscle, contributing to obesity and cognitive impairment — whereas omega-9 fatty acid (oleic acid) may have an opposite effect [JOURNAL OF CLINICAL INVESTIGATION; Benoit,SC; 2577-2589 (2009)]. The figures are based on my best judgement about what I found from various sources, including websites (caveat emptor). Other fat components may not be shown, causing the totals to be less than 100%. The asterisk on coconut oil indicates that it has healthful medium chain triglycerides in the oil. Macadamia oil is 20% palmitolaic acid. Walnuts have health benefits not represented in the oils. Pistachio oil reputedly elevates HDL cholesterol. "<1" indicates "less than one". I have re-arranged the table somewhat based on a de-emphasis of the harm of omega−6s and palmitic acid.
FAT/OIL | % OMEGA−3 | % OMEGA−6 | % PALMITATE | % OMEGA−9 |
|---|---|---|---|---|
| hazelnut oil | 0.1-0.2 | 14.2-13.2 | 4.5-5.9 | 68.8-78.6 |
| canola oil | 9-11 | 21 | 4 | 61 |
| almond oil | <1 | 29 | <1 | 62 |
| chia seed | 60 | 15 | 20 | 5 |
| flax seed | 47 | 25 | 6 | 19 |
| walnut oil* | 1.5 | 60 | <1 | 25 |
| pine nut oil | <1 | 49 | 6 | 24 |
| hemp oil | 25 | 55 | 6 | 12 |
| soybean oil | 7-10 | 51-54 | 10 | 23 |
| coconut oil* | <1 | 15 | 10 | 7 |
| macadamia oil* | 1-2 | 1-3 | 17 | 60 |
| olive oil | 1 | 4-21 | 8-20 | 55-83 |
| avacado oil | <1 | 15 | 15 | 71 |
| cashew oil | <1 | 15 | 10 | 60 |
| pistachio oil* | <1 | 33 | 11 | 69 |
| peanut oil | <1 | 33 | 10 | 47 |
| pecan oil | 1.5 | 37 | 7 | 52 |
| sesame seed oil | <1 | 35-50 | 10 | 35-50 |
| milk fat (butterfat) | 1 | 3 | 31 | 24 |
| pumpkin seed oil | <1 | 20-60 | 13-18 | <1 |
| cacao butter | <1 | <4 | 24-30 | 29-38 |
| sunflower seed oil | <1 | 50-75 | 4-9 | <1 |
| caraway seed oil | <1 | 55-65 | 10-12 | 15-18 |
| grapeseed oil | <1 | 70 | 7 | 16 |
| apricot kernal oil | <1 | 20-35 | 3-6 | 55-70 |
For more dietary oil composition data, click on the names of the carrier base oils at: Carrier — Base Oils
Phospholipids and cholesterol are the principal components of nearly all
cell membranes. The backbone of a phospholipid is the same glycerol molecule
that forms the backbone of triglycerides. But instead of 3 fatty acids attached
to glycerol, a phospholipid consists of 2 fatty acids, a phosphate group
and an alcohol. The
most common alcohols are derived from serine, ethanolamine, choline and
inositol. Thus, the most common phospholipids are
phosphatidylcholine, phospatidylethanolamine,
phosphatidylserine, and phosphatidylinositol.
Phosphatidylcholine is also known as lecithin.
"Commercial lecithin" (which is used as an emulsifying agent in food
processing) is a mixture of phospholipids from eggs, soybeans, nuts, etc.,
with phosphatidylcholine as the major ingredient.
![[Phospholipids in Cell Membranes ]](phlipids.gif)
|
Different cells have different quantities of phospholipid in their membranes. Gray matter in the brain is nearly 70% phospholipid, whereas brain white matter is less than half phospholipid because of high concentrations of glycolipid (sugar-fat). There is much variation in the amounts and kinds of phospholipid in membranes. Brain gray matter is 30% phosphatidylcholine whereas brain white matter is 10% phosphatidylcholine. The inner layer of neuron membranes (and cell membranes in general) primarily contain phosphatidylethanolamine & phosphatidylserine, whereas in the outer layer phosphatidylcholine, phosphatidylinositol & sphingomyelin predominate [CHEMISTRY AND PHYSICS OF LIPIDS; Kronke,Ml 101(1):109-121 (1999)]. With age, erythocytes begin to contain increasing amounts of phosphatidylserine in the outer membrane [Ibid.].
Mitochondrial and endoplasmic reticulum membranes are both 40% phosphatidylcholine. But mitochondrial membranes are also 35% phosphatidylethanolamine, whereas endoplasmic reticulum membrane is about 17% phosphatidylethanolamine.
Cells also vary considerably in the kinds of fatty acids attached to phosphatidic acid. In gray matter cell membranes, the fatty acids in the middle position of phospholipids are composed of carbon chains that are longer and more unsaturated than fatty acids found in the membrane phospholipids of most other cells. Fatty acids that are long and highly unsaturated increase membrane fluidity and functionality, which is why DHA and arachidonic acid are highly concentrated in the phospholipids of neuron synapses. Unsaturated fatty acids are also important for membrane activity at the site of hormone receptors. Insulin resistance in adult-onset diabetes is associated with fewer membrane long-chain unsaturated fatty acids due to impaired desaturase and elongase enzyme function [*16].
The alcohol portion of a phospholipid protrudes away from the
membrane, whereas the two fatty acids jut into the membrane. The
middle fatty acid (in the second position) is usually unsaturated
(like DHA or arachidonic acid) whereas the end fatty acid (in the
first position) is usually saturated (like stearic acid). Each of
the 3 groups attached to the glycerol backbone has a special enzyme
that can separate the group from the backbone.
Phospholipase A1
enzyme attacks the attachment of the first fatty acid,
Phospholipase A2
attacks the attachment of the middle fatty acid and
Phospholipase D attacks the alcohol attachment.
Phospholipase C — which is a major toxin secreted by bacteria —
releases 1,2-DiAcylGlycerol (DAG) along with a phosphoryl base.
![[Phospholipase Enzymes in Cell Membranes ]](membrane.gif)
|
Frequently the unsaturated fatty acid stored in the second position of a cell membrane will be arachidonic acid, EPA or DHA (especially in the neurons). The release of arachidonic acid or EPA from cell membranes by Phospholipase A2 allows the enzymes lipoxygenase and cyclooxygenase to form biologically active eicosanoids like prostaglandins (PGZs, first isolated in prostate gland), thromboxanes (TXZs, first isolated in thrombocytes) and leucotrienes (LTZs, first isolated in leucocytes). These eicosanoids can be compared to hormones, except that unlike hormones they are destroyed by local enzymes within seconds or minutes after formation. This limits the activities of eicosanoids to the area where they were released.
Having cell membranes contain fatty acids that can form the
hormone-like eicosanoids gives the body the capacity to produce quick,
localized action in almost any tissue or organ. The most general need
for rapid, local action is the response to trauma. Therefore, eicosanoids are most often
concerned with clotting, inflammation and the initiation of immune defense.
![[Membrane Phosopholipid Hydrolysis by Phospholipases]](./PLipases.gif)
Some membrane phospholipids, such as the phosphatidylinositols, function to convert activity at cell surface G-protein-coupled receptors into intracellular signals. Hydrolysis by phospholipase C and phospholipase D produce the second messengers (intracellular messengers) DiAcylGlycerol (DAG) [which stimulates Protein Kinase C (PKC)] and Inositol triPhosphate (IP3) [which causes intracellular release of calcium]. Activated PKC concentrates in the plasma membrane where it phosphorylates membrane proteins of receptors and ion-channels to inhibit their function (negative feedback). Nuclear Factor kappaB (NF−κB) activated by PKC binds to DNA promoters & enhancers of inflammatory cytokines, among other genes. IP3 binds to the endoplasmic reticulum, releasing calcium stored in that location. DAG can be further hydrolysed by Phospholipase A2 to release more arachidonic acid. Ethanol increases Phospholipase A2 activity, increasing oxidative stress.
Three categories of PhosphoLipase A2 (PLA2) are recognized: secretory PLA2 (sPLA2), cytoplasmic PLA2 (cPLA2) and Ca2+-independent PLA2 (iPLA2). The cPLA2 is activated by Ca2+ to a much greater extent than sPLA2. Whereas sPLA2 is more prominent in inflammatory disease, cPLA2 is more associated with oxidative free radical damage. The cPLA2 shows a marked preference for hydrolyzing oxidized arachidonic acid in cell membranes, which may be important for membrane maintenance when sufficient ATP is available to synthesize and insert fresh arachidonic acid into membranes. Arachidonic acid is a particularly common constituent of brain neuron membranes and the massive release of arachidonic acid in cerebral cortex ischemia/reperfusion plays a significant role in exacerbating ischemic/reperfusion damage. The activity of cPLA2 on inner mitochondrial membranes can also be exacerbated under conditions of high oxidative stress.
There are three series of the prostaglandin and thromboxane
eicosanoids: one derived from DGLA (series 1), one derived from arachidonic
acid (series 2) and one derived from EPA (series 3). The series number
indicates the number of double-bonds in the prostaglandin or thromboxane.
Series 1 prostaglandins are not as common in the body as
series 2 or 3 — in part because DGLA is not plentiful in many tissues.
Series 2 predominates over series 3 for at least three
reasons: (1) arachidonic
acid is more readily released from cell membranes than EPA, (2) arachidonic
acid reacts far more avidly with cyclo-oxygenase enzyme than does EPA and
(3) contemporary Western diets contain large amount of linoleic acid, which
results in large amounts of arachidonic acid in cell membranes. The
predominance of arachidonic acid eicosanoids over EPA-eicosanoids due to
excessively high dietary omega−6 (compared to omega−3) is at the root of
many modern health problems, only some of which are concerned with the immune
system.
![[ Eicosanoid Abbreviations ]](eicosan.gif)
|
![[Series-2 Eicosanoids (from Arachidonic Acid) ]](cycloox1.gif)
|
![[Series-3 Eicosanoids (from EPA) ]](cycloox2.gif)
|
Cyclooxygenases (COXs) can be divided into COX-1 (present at constant levels in most cells & tissues) and COX-2 (normally absent from most cells, but rapidly rising to high levels in response to growth factors, cytokines, hypoxia, toxins and other stimuli). COX-1 is regarded as a "housekeeping" enzyme responsible for such functions as maintaining gastrointestinal mucosal integrity and regulation of kidney blood flow. COX-2 expression in response to inflammation & mitogens may lead to cancer. COX-2 enhances the formation of prostaglandins that mediate pain & inflammation. The attempts to create drugs that specifically inhibit COX-2 without inhibiting COX-1 (which can result in gastrointestinal bleeding) is based on this simplification [BIOCHEMICAL PHARMACOLOGY 65:153-159 (2003)]. The gamma-tocopherol, but not alpha-tocopherol, form of Vitamin E has been shown to be anti-inflammatory by inhibition of the COX-2 enzyme (independent of antioxidant activity) [PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES (USA); Jiang,Q; 97(21):11494-11499 (2000)].
Unlike COXs, which are active in most body cells, lipogenase enzyme is primarily
active in cells of the immune system. Lipogenase creates hydroperoxides from polyunsaturated
fatty acids by an insertion of molecular oxygen.
Both arachidonic acid (omega−6) and EPA (omega−3) can be acted-on by
lipoxygenase enzyme, rather than by cyclooxygenase — resulting in
leucotrienes (LTZs) rather than the prostaglandins (PGZs)
& thromboxanes (TXZs) produced by cycloxoygenase. Lipoxygenase produces
series 4 leucotrienes from arachidonic acid or series 5 leucotrienes from
EicosaPentoic Acid (EPA). Lipoxygenase is primarily found in granulocytes.
HydroxyPeroxyEicosaTetraEnoates (HPETEs)
are reactive hydroperoxides which can contribute to cellular damage in
atherosclerosis [THE JOURNAL OF CLINICAL INVESTIGATION 111(8):1107-1113(2003)].
HPETEs like other organic hydroperoxides (ROOH) can lead to lipid peroxidation
by Fenton-like reactions.
![[ Arachidonic Acid conversion
to Leukotrienes via HPETEs]](Lipoxygenase.jpg)
|
|
Arachidonic acid oxidized in the membranes of neurons which has been subsequently
liberated by phospholipase results in enzymatic products known as isoeicosanoids
because they are isomers of normal eicosanoids. Isoprostanes (isomers of
cyclooxygenase-derived prostaglandins) and particularly F2-isoprostanes
(isomers of prostaglandin F2α) have attracted special interest because they
are chemically stable and can be sensitively measured using mass spectrometry.
F2-isoprostanes have been used to quantify lipid peroxidation in atherosclerotic
plaques, in the brains of Alzheimer's
Disease patients and in the urine of smokers & myocardial infarction patients.
Elevated levels of F2α in pericaridial fluid is correlated with severity of heart
failure [CIRCULATION; Mallat,Z; 97(16):1536-1539 (1998)].
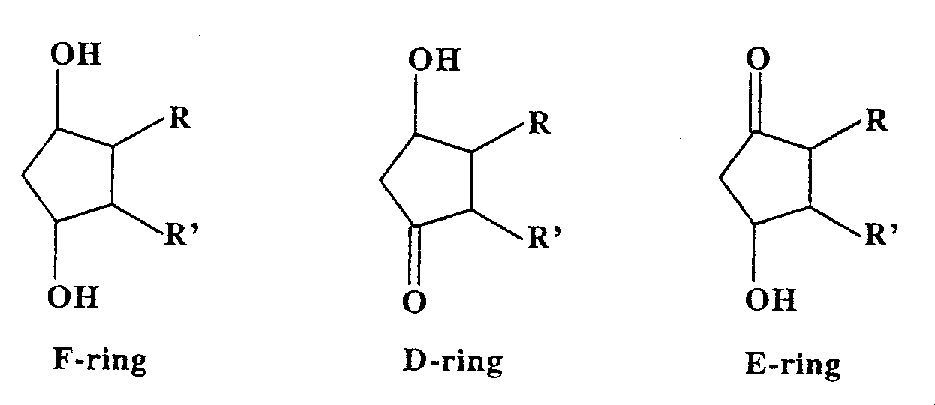
Analogous F-ring oxidized isomers of DHA breakdown products known as
F4-neuroprostanes have been isolated in elevated quantity in both
cerebrospinal fluid and brain tissue of Alzheimer's Disease patients [CELLULAR
AND MOLECULAR LIFE SCIENCES 59:808-820 (2002)].
| Normal F-Ring Prostaglandin | F2-isoprostanes |
|---|---|
![[Normal F-Ring Prostaglandin]](PGF2alpha.gif)
|
![[F<SUB>2</SUB>-isoprostanes]](F2isoPs.gif)
|
Cell membrane enzyme activity is significantly influenced by membrane fatty acid composition. Rats fed unsaturated fats show higher sodium-potassium pump activity than rats fed saturated fats [NEUROCHEMISTRY INTERNATIONAL; Srinivasarao,P; 31(6):789-794 (1997)]. Increasing membrane DHA content is a mechanism for cold adaptation in fish. But the longer lifespan of birds compared to similar-sized mammals is attributed to greater saturated fat content in membranes (lower peroxidizability) [JOURNAL OF EXPERIMENTAL BIOLOGY; Hulbert,AJ; 206(Pt 14):2303-2311 (2003)].
One very general way of classifying immunity is to distinguish between acquired immunity and innate immunity. Both systems make use of phagocytes (cells that "eat" foreign substances) and antibodies (Y-shaped chains of proteins known as immunoglobulins). Acquired immunity is seen when the immune system has identified a foreign protein as being an antigen. For innate immunity, however, there is a generalized reaction against tissue trauma and infectious agents that could as easily be described as inflammation as immunity. The swelling of inflammation is associated with immunoglobulin-E (IgE) and with an increased blood vessel permeability which allows Natural Killer cells, macrophages and neutrophils to leave the blood stream, migrate to the injured area and engulf the invaders. Although innate immune defense is not very specific, it is rapid. By contrast, the immune cells (lymphocytes) and immunoglobulins (IgM and IgG) of acquired immunity are slower to develop, but can be very powerful and very specific against antigens.
|
The eicosanoids produced from arachidonic acid cause a stronger inflammatory response than the eicosanoids from EPA or DGLA. The arachidonic acid products LeukoTriene B4 (LTB4) and ProstaGlandin E2 (PGE2) are powerful promoters of inflammation. LTB4 is only formed in granulocytes subject to stimuli favoring action by 5-lipoxygenase activating protein [ARCHIVES OF BIOCHEMISTRY AND PHYSICS 356(1):71-76 (1998)]. Both PGE2 & LTB4 increases tissue swelling (edema) by making the vascular endothelium more leaky. LTB4 stimulates superoxide production by neutrophils, activates Natural Killer cells and powerfully attracts inflammatory leukocytes. PGE2 increases sensitivity to pain, raises temperature and increases the formation of the allergic antibody Immunoglobulin E (IgE) [*17]. By promoting the production of less inflammatory eicosanoids than arachidonic acid, EPA is anti-inflammatory. A significant inverse exponential relationship has been demonstrated between human EPA supplementation and synthesis of the pro-inflammatory cytokines TNF−α and IL−1β [AMERICAN JOURNAL OF CLINICAL NUTRITION; Caughey,GE; 63(1):116-122 (1996)]. Moreover, EPA has been shown to facilitate removal of inflammatory cytokines [CHEMISTRY & BIOLOGY; Tjonahen,E; 13(11):1193-1202 (2006)].
Aspirin irreversibly blocks the enzyme cyclo-oxygenase, which prevents arachidonic acid from being converted to prostaglandin — thereby limiting inflammation and pain. Gamma-tocopherol (the major form of Vitamin E in food, in contrast to alpha-tocopherol, which is the major form of Vitamin E in supplement pills) also blocks cyclo-oxygenase and reduces proinflammatory PGE2 & LTB4 formation [FASEB JOURNAL 17:816-822 (2003)].
Elevated body temperature, increased sensitivity to pain, inflammation and allergic reactions are important body defense mechanisms. Inflammation reduces the spread of infection. Elevated body temperature can kill bacteria and viruses. Increased sensitivity to pain reduces movement of injured body parts, preventing injury from getting worse. But when taken to an extreme, inflammatory reactions can result in auto-immune disease, septic shock, asthma and even fatal anaphylactic shock. Chronic inflammation of the pancreas, colon and other organs increase the risk of cancer in those organs. The increasing incidence of allergies and seasonal asthmas in modern society has been attributed to increased levels of the arachidonic acid eicosanoid LTB4 due to excessively high dietary omega−6 intake relative to omega−3 [*18].
Gamma-Linolenic Acid (GLA) has been shown to be effective against the inflammation of rheumatoid arthritis in a number of studies. Although one might expect that GLA could lead to the formation of arachidonic acid's pro-inflammatory eicosanoids, there is instead a production of the anti-inflammatory prostaglandin PGE1 of the 1−series [*19,*20]. It may be that rheumatoid arthritis patients suffer from impaired function of desaturase enzymes, preventing arachidonic acid formation. In ulcerative colitis, an inflammatory condition in which desaturase enzymes are normal, both omega−3 oils from fish and perilla have been used for treatment [*21].
Feeding laboratory animals diets rich in omega−3 fatty acids (linseed or fish oil) reduces Natural Killer cell and cytotoxic T−lymphocyte activity [*22], but stimulates the more antigen-specific immunoglobulins IgM and IgG [*17]. Innate immune response, although closely tied to inflammation can be separated from inflammation to some extent. One experiment showed that both fish oil and safflower oil reduced the secretion of Interleukin-6 (a cytokine that activates lymphocyte immune-cells and increases antibody production), but that only fish oil inhibited the secretion of Tumor Necrosis Factor alpha (a cytokine that increases fever, shock and blood vessel permeability) [*23].
As a first-line defense against infection, the inflammatory/innate-immunity response can sometimes mean the difference between life and death. Adding fish oil (which contains both DHA and EPA) to the diet of rats increased the likelihood that the rats would die when subjected to bacterial infection [*24]. Mixtures of DHA and EPA strongly reduce Natural Killer and Lymphocyte Activated cells [*25]. But immune suppression appears to be more due to EPA than to DHA [*22]. In fact, when total fat intake is low, DHA shows no inhibition of immune system function, even though about 9% of DHA is converted back to EPA [*26]. The omega−6 to omega−3 proportions may also determine immune suppression. An experiment, using a mixture of safflower oil and fish oil showed no immunosuppressive effect in rats when the omega−6 to omega−3 ratio was approximately 2−to−1 [*23].
Cells of the immune system (like T-Cells, B-Cells and Macrophages) have membranes that are particularly rich in long-chain unsaturated fatty acids (such as arachidonic acid or EPA). Unlike prostaglandins, which are formed from most cells of the body, leukotrienes are made predominantly by polymorphonucleocytic leucocytes, macrophages and mast cells.
The G protein-coupled receptor GPR120 is highly expressed in adipose tissue and pro-inflammatory macrophages, functioning as an omega-3 fatty acid sensor. Through action on GPR120, treatment with both DHA and EPA was shown to reduce inflammatory cytokines while increasing insulin sensitivity [CELL; Oh,DY; 142(5):687-698 (2010)]. Chronic inflammation is an important source of insulin resistance in obesity.
Because immune system cells are so rich in arachidonic acid and EPA (and also because of their mobility and functions) immune system cells are more vulnerable to free-radical oxidation than other cells. The nutrients that most profoundly improve immune function are vitamin C, vitamin E, selenium, glutathione and zinc [*27]. All of these nutrients are antioxidants, although zinc's effects are more due to direct actions on immune cell function than due to its anti-oxidant properties [*28]. Vitamin E opposes some, but not all of the increased lipid peroxidation and immune suppression seen in essential fatty acid supplementation [*29]. Vitamin E promotes immune function by reducing PGE2 synthesis and thus increasing T-cell proliferation, IL-2 production and antibody production, while reducing IL-6 production. By scavenging the hydroperoxide necessary for COX activity, Vitamin E opposes the increase in PGE2 formation that is typically seen in aging [AMERICAN JOURNAL OF PHYSIOLOGY; Wu,D; 275(3 Pt 1):C661-C668 (1998)]. PGE2 is known to suppress lymphocyte proliferation, to suppress synthesis of chemical factors (lymphokines) influencing the immune system and to contribute to the auto-immune diseases that increase with aging [*30].
High fat diets are well-known to be associated with certain kinds of cancers, including breast cancer, in particular [*31]. Although butterfat stimulates breast cancer when compared with a fat-free diet, safflower oil margarine (linoleic acid, an omega−6) has been shown to induce breast cancer much more strongly [*6]. Linoleic acid is the fat that most frequently is associated with cancer, whereas omega−3 fatty acids like DHA and perilla-oil suppress cancer [*32,*33]. It has been theorized that linoleic acid causes cancer by chronic overproduction of the inflammatory arachidonic acid eicosanoids, which stimulate the proliferation of mutated cells [*6]. Omega−3 fatty acids reduce cancer risk by markedly inhibiting Activator Protein 1 (AP−1), a transcription factor which promotes cancerous proliferation and metastasis. Omega−6 fatty acids promote cancer by blocking omega−3 activity [PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES (USA); Liu,G; 98(13): 7510-7515 (2001)].
PGE2 resulting from COX-2 enzyme is found in high levels in colorectal carcinomas where the prostaglandin promotes growth through transactivation of epithelial growth factor receptor signalling. PGE2 also induces expression of Vascular Endothelial Growth Factor (VEGF) in colon cancer cells, resulting in the neoangiogenesis (new blood vessels) required by growing tumors [JOURNAL OF CLINICAL INVESTIGATION; Williams,CS; 105(11):1589-1594 (2000)]. Nitric oxide is a regulator of COX-2 expression in the colon, and the phytochemical curcumin (an inhibitor of inducible nitric oxide synthetase found in curry) has been shown to reduce pre-cancerous colon lesions by 45% [CARCINOGENESIS; Rao,CV; 20(4):641-644 (1999)].
Although too much linoleic acid can increase the risk of cancer, a form of linoleic acid in which the double-bonds are closer together — known as Conjugated Linoleic Acid (CLA) — actually reduces cancer risk. CLA has been shown to significantly inhibit prostate cancer proliferation [*34] and breast cancer formation [*35] in experimental animals. Large quantities of CLA are not needed since the maximum anti-cancer effect is seen when CLA is no more than 1% of calories. There is controversy over whether CLA is an anti-oxidant. Some researchers who believe that CLA is not an anti-oxidant believe that CLA's effects are due to its blockage of arachidonic acid formation [*36,*37]. CLA has also been shown to reduce atherosclerosis in rabbits [*38]. The best dietary sources of CLA are the food products of ruminant animals (ie, animals that "ruminate" their food by chewing a cud, like cows or deer), such as beef, milk, yoghurt and cheese.
For technical details about the nature of cancer (and methods of prevention) — see my essay Cancer.
Insulin stimulates the delta−6−desaturase enzyme. Therefore, this enzyme's activity is much reduced in diabetes. Reduced delta-6-desaturase activity affects the omega−6 products significantly more than the omega−3 products [*39]. Although excessive arachidonic acid is associated with many modern illnesses, diabetics suffer from the effects of too little arachidonic acid. Arachidonic acid is essential for leucocyte (white blood cell) function. Reduced leucocyte function makes diabetics more vulnerable to infection [*40].
Arachidonic acid deficiency in diabetics also reduces the activity of the "sodium pump", slowing nerve conduction velocity and ultimately leading to neuropathy (diseased nerves) [*41]. Insulin is the best treatment for these arachidonic acid deficiency conditions, but gamma-linolenic acid (GLA) also restores function. GLA bypasses the delta-6-desaturase step in arachidonic acid synthesis. Evening primrose oil is the most effective omega−6 treatment for diabetic neuropathy, despite the fact that borage oil has more GLA. Although fish oil can be somewhat beneficial when used alone, fish oil can detract from the effectiveness of evening primrose oil when they are used in combination [*42].
Alcoholics suffer from disturbances of fat metabolism, notably in the liver. The liver is the most active site of delta-6-desaturase activity in the body. In fact, many (if not most) cells in the body have no delta-6-desaturase enzymes and are dependent upon the liver for omega−3/omega−6 desaturase/elongase products. Neuropathy and other conditions resulting from desaturase dysfunction in alcoholics are benefitted by both evening primrose oil and fish oil in combination [*43]. Arachidonic acid deficiency is the most serious problem for alcoholics, however, so evening primrose oil seems to be the best therapy [*44].
DHA and arachidonic acid are the predominant essential fatty acids in the human brain. Neurons cannot synthesize arachidonic acid, but astrocytes and cerebral epithelial cells have enzymes that can. Membrane fluidity and saturated fatty acid content of brain cell membranes decline with age [MOLECULAR AND CELLULAR BIOCHEMISTRY; Pu,L; 198(1-2):69-78 (1999)]. The ability of enzymes to produce the omega−6 and omega−3 family of products of linoleic and alpha-linolenic acid declines with age. One experiment showed that desaturase enzyme function in old rats was only 44% of the desaturase function in young rats [*39]. This decline in desaturase activity has provided a rationale for supplementation with GLA (bypassing the delta-6-desaturase enzyme), and for supplementation with the omega−3 products EPA and DHA. Fatty acids in human gray matter phosphatidylethanolamine is roughly 25% DHA, 25% stearic acid, 14% arachidonic and 12% oleic acid. In the outer segments of retina photo-receptors of the eye, DHA accounts for more than 50% of the fatty acid content, probably because of the high membrane fluidity required for sensitivity to light.
In the last third of pregnancy, and in the first four months after birth, rapid brain growth in the human infant requires large amounts of omega−3 and omega−6 essential fatty acids. Human milk contains (in total fatty acids by weight) 12% linoleic acid, 0.5% alpha-linolenic acid, 0.6% arachidonic acid and 0.3% DHA [*45]. Infant formulas frequently have not contained arachidonic acid or DHA. One study showed that by (or just before) age 8, children who had been breast-fed as infants had an 8.3-point IQ advantage over children who had received formula [*46]. The study corrected for the education and social class of the mother.
Support for the idea that DHA is critical for brain development came from an experiment which studied the effects of adding DHA (in the form of fish oil) to infant formula. At both 16 and 30 weeks of age the breast-fed and supplement-formula-fed infants showed significantly better visual acuity than the placebo-formula-fed infants [*47]. Arachidonic acid supplementation is also needed because DHA supplementation given alone lowers arachidonic acid levels [*48] and because arachidonic acid is essential for growth [*49,*50]. The lipoxygenase eicosanoids of arachidonic acid contribute to the ability of Nerve Growth Factor (NGF) to increase neurite outgrowth [*51]. Deficiency of arachidonic acid during brain development is less reversible than deficiency of DHA [*6]. Evidently an infant's desaturase and elongase enzymes are not fully developed because no amount of alpha-linolenic acid supplement can provide enough DHA for neural development [*52]. More recent reviews have firmly recommended the inclusion of arachidonic acid and DHA in the formula of premature babies [*53].
Children with developmental coordination disorder showed improved reading, spelling, and behavior when supplemented with omega-3 and omega-6 fatty acids [PEDIATRICS; Richardson,AJ; 115(5):1360-1366 (2003)].
It seems reasonable to wonder if dietary fat or fat supplements affect the mental function of adults. Experiments on rats indicate that manipulation of dietary fats can alter the fatty acid composition of brain-cell membranes — with effects on behavior. A diet high in saturated fat was shown to "impair a wide range of learning and memory functions" [*54]. Soybean oil has more alpha-linolenic acid than sunflower oil. Soybean-fed rats have shown significantly better learning and less sensitivity to pain than safflower-fed rats [*55]. Recent experiments testing relative concentrations of linoleic acid to alpha-linolenic acid in the range of from 3-to-1 to 6-to-1 verified that a 4-to-1 ratio is optimal for spatial learning and pain tolerance in rats [*56]. EPA & DHA supplementation increases neurogenesis in old rats [JOURNAL OF NEUROSCIENCE RESEARCH; Dyall,SC; 88(10):2091-2102 (2010)].
The influence of essential fatty acid supplement on brain membrane content declines with maturity, however. Adult cell membrane content of DHA and arachidonic acid is only slowly altered by diet or supplement. If dietary intake of essential fatty acid is low, the body will sacrifice essential fatty acid content of cell membranes outside the brain before neurons are affected. A study of elderly men, however, showed greater cognitive impairment among those with a history of high dietary linoleic acid compared with controls, whereas those with high fish consumption showed reduced cognitive decline [*57]. Another study showed reduced cognitive decline in elderly men consuming EPA & DHA [AMERICAN JOURNAL OF CLINICAL NUTRITION; van Gelder,BM; 85(4):1142-1147 (2007)]. One experiment showed promotion of neuron growth by DHA and inhibition by arachidonic acid [*58]. But an experiment on adult mice showed that excessively high supplements of DHA without arachidonic acid supplementation can impair physical and cognitive performance [*59]. Arachidonic acid may facilitate LTP-type learning in the hippocampus of the brain [*51].
An experiment studying maze-learning in rats demonstrated that, after training, the rats showed less cholesterol and more membrane fluidity in the hippocampal and cortical regions of the brain [*60]. Adult mice fed fish oil for 12 months showed more brain DHA, less brain arachidonic acid, more synaptic membrane fluidity and higher maze-learning ability [*61]. Rats fed perilla oil showed 30% more hippocampal neuron synaptic vesicle density and improved learning compared to rats fed safflower oil [*62].
Measurements of fatty acid content of brain cell (neuron) membranes show decreased DHA with aging. Since DHA is particularly concentrated in synaptic membranes, lowered DHA levels may contribute to declining brain function. DHA is also reduced when the brains of rats are experimentally exposed to high oxygen levels — suggesting that free-radical oxidation is causing the depletion in both cases. Vitamin E treatment protected the rats from neuron damage from the oxygen. This suggests that Vitamin E may be important for prevention of neurodegeneration in humans [*63]. (For more detail about DHA — see my essay DHA for Hearts and Minds).
Arachidonic acid can be released from cell membranes exposed to neurotransmitters, neuromodulators and neurohormones. The release of arachidonic acid can directly modify neuron excitability by binding with hydrophobic binding sites of ion channels or can be metabolized to eicosanoids that ineract with neuronal eicosanoid receptors to have a second messenger effect. Treatment of the elderly with both DHA and arachidonic acid (ARA) has been shown to increase both cognitive function and coronary microcirculation [JOURNAL OF PHARMACEUTICAL SCIENCES; Kiso,Y; 115(4):471-475 (2011)].
Arachidonic acid is known to worsen brain damage during stroke and other conditions of oxygen depletion. Although Vitamin E reduces the toxic effects of arachidonic acid in hypoxic conditions, N-Acetyl-Cysteine (NAC, a nutrient which increases glutathione synthesis) completely blocks the arachidonic acid toxicity. Because NAC is not an effective anti-oxidant against superoxide, the arachidonic acid toxicity must be due to lipid peroxidation [*51].
Since the 1950s it has been believed that schizophrenia is caused by brain disturbances involving the neurotransmitter dopamine. Recently it has been observed that many schizophrenics have reduced levels of DHA and arachidonic acid — and a sizable proportion of these patients do not flush red on 200 mg doses of niacin. DHA is known to be highly concentrated in synapses, suggesting that reduced sensitivity to dopamine due to low levels of DHA and arachidonic acid in dopamine receptors may be a more fundamental cause of schizophrenia than dopamine deficiency [*64]. Fish oil (EPA & DHA) has shown to be of value in reducing bipolar symptoms, but EPA or DHA alone has not [ARCHIVES OF PSYCHIATRIC NURSING; Turnbull,T; 22(5):306-311 (2008)].
As brain cell membranes age, the ratio of cholesterol to phospholipid increases and membrane fluidity decreases. A similar effect is seen in brains which are becoming tolerant to ethyl alcohol (beverage alcohol). Experiments exposing rats to ethyl alcohol showed tissue depletion of DHA and arachidonic acid, particularly in the liver [*65,*66]. Phosphatidylethanolamine in the cerebral cortex gray matter of the brains of alcoholics show lower levels of DHA and arachidonic acid [*67]. Methionine (or S-adenosylmethionine, SAM) has reversed the DHA and arachidonic acid depletion in the liver following alcohol treatment of rats [*66]. Oxidized DHA and arachidonic acid enzyme products (isoprostanes and neuroprostanes) are both markedly eleveated in the cerebrospinal fluid of Alzheimer's Disease patients as compared to age-matched controls. Lipid peroxidation would be an expected consequence of inflammatory processes associated with Alzheimer's Disease, but could also be indicative of oxidation as a cause of the disease.
The low death rate from coronary heart disease among Greenland Eskimos led scientists to suspect that high fish consumption might be protective. A 20-year study of 852 middle-age Dutch men showed that coronary artery disease was more than 50% lower among the men who consumed at least 30 grams of fish per week, when compared with men who did not eat fish [*68]. A 30-year study of over 2,100 Chicago men showed a 62% risk of coronary heart disease and 56% risk of sudden myocardial infarction for men who ate at least 35 grams of fish daily, compared to those who ate none [*69].
Series 3 prostaglandins & thromboxanes inhibit the release of arachidonic acid from phospholipids and thus reduce formation of series 2 prostaglandins & thromboxanes. PGI3 is as potent an antiaggregator as PGI2, whereas TXA3 is a weaker platelet aggregator than TXA2 — so the series 3 products result in a less net clotting. TXA2 causes potent vasoconstriction.
Oxidative stress is known to contribute to atherosclerosis, but it is usually attributed to oxidation of LDL-cholesterol, causing it to adhere to artery walls. Some oxidative effect is also due to vascular smooth muscle cell proliferation induced by arachidonic acid derived eicosanoids functioning as transcription factors. The arachidonic acid lipoxygenase metabolites 12-HPETE and 15-HPETE act as mitogens by increasing AP-1 (Activator Protein-1) transcription factor activity [JOURNAL OF BIOLOGICAL CHEMISTRY; Rao,GN; 271(44):27760-27764 (1996)].
Fish oil has been shown to lower LDL-cholesterol by about 13% [*70], to lower blood pressure [*71], and to dramatically lower blood triglycerides [*72,*73]. The effect on triglycerides, in particular, appears to be due to EPA and DHA, because flaxseed (ie, linseed oil, which is over 50% alpha-linolenic acid) did not lower triglycerides [AMERICAN JOURNAL OF CLINICAL NUTRITION; Harris,WS; 65(Suppl 5):1645S-1654S (1997)]. Other analysis has determined that DHA alone has no effect on blood triglycerides, but that EPA alone is capable of lowering blood triglycerides by about 30% [*74]. Purified DHA was shown to lower blood pressure and reduce blood viscosity. The evidence indicated that DHA increases red blood cell membrane fluidity, thereby increasing the deformability of the blood cells so that they can move through capillaries more easily and thus lower blood viscosity and blood pressure [*75]. Fish oil has been shown to inhibit atherosclerosis in monkeys fed an atherogenic diet [ATHEROSCLEROSIS, THROMBOSIS, AND VASCULAR BIOLOGY; Davis,HR; 7(5):441-449 (1987)].
High fasting blood insulin concentrations has been determined
to be an independent risk factor for ischemic heart disease [*76].
The fatty acid content of muscle membranes is critical in determining
insulin sensitivity. A high-fat diet can increase
insulin resistance,
but one study showed that both dietary omega−3 and
omega−6 fatty acids can increase membrane
fluidity and thereby improve insulin sensitivity [*77]. A study
on rats, however, showed that omega-6 fatty acids severely
increased insulin resistance, whereas omega-3 fatty acids had the
opposite effect [DIABETES; Storlein,LH; 40(2):280-289 (1991)].
A human study showed fish oil both increases insulin resistance
and reduces pro-inflammatory C-reactive
protein [HORMONE AND METABOLIC RESEARCH; Tsitouras,PD;
40(3):199-205 (2008)]. Long-chain fatty acids incorporated
in cell membranes increases membrane fluidity and increases both
the number and efficiency of insulin
receptors [PROSTAGLANDINS, LEUKOTRIENES, AND ESSENTIAL FATTY ACIDS;
Das,UN; 72(5):343-350 (2005)].

The protective effect of fish oil against cardiac arrhythmias (irregular heartbeats) has been strikingly illustrated by two similar experiments, one performed on rats [*78] and the other on marmoset monkeys [*79]. Middle-aged animals were fed sheep fat (saturated fat), sunflower seed oil (omega−6) or fish oil (omega−3) for 12 weeks (for rats) or for 24-30 months (for monkeys). With both rats and monkeys arrhythmia was produced in over 40% of the animals fed sheep fat, roughly 10% of the animals fed safflower oil and in none of the animals who were fed fish oil.
Although human epidemiological studies and clinical trials on the use of fish oil (DHA & EPA) have shown significant reduction in cardiovascular deaths, the results for arrhythmias are mixed. One large review showed no benefit against arrhythmia [BMJ; Leon,H; 337:a2931 (2008)], whereas a similarly large review showed some benefit against sudden cardiac death (arrhythmia) [CLINICAL CARDIOLOGY; Marik,PE; 32(7)365-372 (2009)]. Another review noted that fish oil is anti-arrhythmic for patients with prior myocardial infarction, but fish oil may be pro-arrhythmic in patients with re-entrant arrhythmia (due to acute ischemia or chronic ventricular fibrillation) [CARDIOVASCULAR RESEARCH; Den Ruijter,HM; 73(2):316-325 (2007)]. A study of diabetic patients who had suffered a myocardial infarction showed that low-dose supplementation with omega-3 fatty acids had a protective effect against ventricular arrhythmia [DIABETES CARE; Fromhout,D; 34(12):2515-2520 (2011)]. Fish oil has been shown to reduce heart rate and improve heart rate variability in overweight, sedentary persons [BRITISH JOURNAL OF NUTRITION; Ninio,DM; 100(5:1097-1103 (2008)].
Phosphatidylethanolamine from monkey heart tissue showed 5 times more (over 25% total) DHA in the fish-oil fed monkeys than in the other two groups. EPA accounted for over 6% of the fatty acid phosphatidylethanolamine of fish-oil fed monkeys, and was undetectable in the other two groups. A similar experiment on rats using purified DHA and purified EPA, rather than fish-oil, indicated that DHA is responsible for most of the anti-arrhythmic effect [*80]. It is the DHA release from membrane breakdown, rather than DHA in the bloodstream, which is protective [*81]. Moreover, DHA in the membrane increases the efficiency of the heart cyclic-AMP (a cell messenger molecule) [*82].
Although most fish oils are high in EPA and DHA, there are some fish oils which are not. Flounder, swordfish and sole are particularly low in EPA and DHA. Fish oils having the highest levels of EPA and DHA include mackerel, herring and salmon. Some fish, such as cod and haddock, store most of their fat in the liver, therefore the liver oils of these fish should be taken rather than the fillet.
Increased fish oil consumption, however, is associated with increased lipid peroxidation in heart, liver and lung tissue — moreso than in the brain [*83]. Again, Vitamin E has been recommended for those who have a high fish oil consumption to reduce lipid peroxidation [*84]. In some cases, consumption of fish can harmful due to high levels of mercury (for more detail about mercury risk — see my essay Is Mercury in Fish a Health Hazard?). Lipid peroxidation of LDL cholesterol is believed to initiate arterial wall injury and facilitate the formation of atherosclerotic foam cells. Leucocytes & endothelial cells may contribute to atherosclerosis through inflammation induced by 5-lipoxygenase [PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES (USA) 100(3):1238-1248 (2003)].
(For more details concerning cardiovascular disease, risk factors and prevention — see my essays Sudden Cardiovascular Death and Prevention of Cardiovascular Disease.)
In the face of all the data, there remains the question "What omega−3 and omega−6 fatty acids should be included in the diet, and in what quantities & proportions?" One standard which is not of much use in this case is the amount of essential fatty acid intake below which a deficiency disease occurs. It is very difficult to produce a deficiency disease of essential fatty acid in normal adults, even in the poorest of diets. The important question is "What levels of essential fatty acids provide optimum health?" This question still results in considerable controversy among nutritionists.
Deficiency disease results from diets where linoleic acid is less than 1% of total calories. Governmental agencies of various nations (including the World Health Organization) recommend an omega−6 intake of 1-3% of total calories, increased to 3-6% during pregnancy and lactation. Since deficiency disease symptoms have not been unquestionably demonstrated for the omega−3 fatty acids (DHA, EPA and alpha-linolenic acid) there is controversy about how much to recommend or whether to recommend omega−3 supplementation at all [*85]. Ironically, the avidity with which enzymes preferentially utilize omega−3 fatty acids could misleadingly make it appear that omega−6 is more essential. When stated, omega−3 recommendations are often given in relation to omega−6 intake, due to the enzyme competition between the two fatty acid families.
The same enzymes that produce the omega−6 products from linoleic acid also operate to produce the omega−3 products from alpha-linolenic acid. The competition for these enzymes by omega−6 and omega−3 fatty acids is unequal, however. Alpha-linolenic acid at 0.5% of calories can suppress arachidonic acid production to the same degree as linoleic acid at 7% of calories can suppress DHA production. Nonetheless, a 14-to-1 ratio of omega−6 to omega−3 is not necessarily optimal.
A 4-to-1 ratio has been cited as optimal in a brain function experiment [*56], whereas another experiment showed 2-to-1 is the best ratio for immune function [*23]. A 5-to-1 ratio has been recommended on the basis of the ratio of omega−6 to omega−3 in human milk. But 5-to-1 is probably just a reflection of the dietary intake of contemporary mothers. The content of DHA and EPA in human milk has been increased experimentally by giving fish oil supplements to lactating women [*86].
Some advocates of "a return to natural levels" of fatty acid intake recommend dietary ratios of 1-to-1. Up until about 200 years ago, the human diet contained much higher levels of omega−3 fatty acids. The meat of wild animals that forage for food is rich in EPA, unlike the meat of domesticated animals that have been fattened with grains. High levels of dietary omega−6 vegetable oils are a modern phenomenon. A higher ratio of omega-3/omega-6 is associated with higher bone mineral density in older human adults [AMERICAN JOURNAL OF CLINICAL NUTRITION; Weiss,LA; 81(4):934-938 (2005)]. EPA competes with arachidonic acid for the cyclo-oxygenase enzyme, reducing the production of thromboxane A2, the most powerful platelet aggregating agent known. Some people feel that restoration of the historic omega−3/omega−6 ratios is a more "natural" way of preventing heart attack than using aspirin to irreversibly inactivate cyclo-oxygenase [*15]. Aspirin, even in normal therapeutic doses, can produce dizziness, migraine headaches, depression, anxiety, and stomach irritation or stomach bleeding [*87].
The high levels of omega−6 fatty acids in modern diets may even adversely affect omega−3 utilization by cellular mechanisms that reduce desaturase formation [*88]. Rats on a perilla oil diet which results in omega−6 to omega−3 ratios of approximately 1-to-4 showed the longest lifespans. Eskimos have low heart and autoimmune disease on a fish oil diet that gives a 1-to-3 ratio [*6]. Concern that the anti-clotting effect of fish oil might lead to increased incidence of stroke are apparently unfounded, since stroke is primarily the result of high blood pressure and weakened blood vessels. Nonetheless, dogs fed a relatively high omega-3/omega-6 ratio diet showed increased lipid peroxidation and reduced plasma alpha−tocopherol along with depressed PGE2 and immune response [JOURNAL OF NUTRITION; Wander,RC; 127(6):1198-1205 (1997)].
It would seem prudent to reduce fat intake of nonessential fats to as low a quantity as possible, so long as absorption of oil-soluble vitamins is not impaired. Dietary fat could mainly consist of essential fats. The value of EPA or DHA to prevent serious heart problems should be enough to encourage anyone to take an amount of these nutrients up to 1% of total calories. For a person on a 2,500 calorie daily diet, this would mean 5 to 7.5 grams of essential fatty acids per day with perhaps 500 mg each of EPA and DHA. But too much EPA and DHA can be harmful. When fish oil supplies up to 12% of calories, both the brain and the liver show arachidonic acid deficiencies [*48]. The same effect would probably be seen with excessive amounts of other omega−3 supplements, like linseed or perilla oil.
It would be nice to have a single essential fatty acid formulation that optimizes benefits to the heart, the brain and the immune system for people of all ages, but this is probably unrealistic. Since DHA is responsible for most of the omega−3 benefits to the brain and the omega−3 prevention of heart arrhythmia — while avoiding most of the immune-system depression due to EPA — omega−3 formulations that are higher in DHA rather than EPA should be preferred. For people who dislike fish oil, especially young people with healthy immune systems and functioning desaturase enzymes, perilla oil or linseed oil may be adequate to obtain a better balance between omega−6 and omega−3. Nonetheless, only about a fifth of alpha-linolenic acid is normally converted to DHA and EPA, partially because the body more readily burns unsaturated fats for energy than saturated fats [*6].
But for the very elderly, immune function might be the paramount consideration. Biomarkers of immune function were shown to be a very good predictor of the 2-year survival of 102 elderly people between the ages of 86 and 92 [*89]. A normally mild disease can be fatal to a person with a weakened immune system. One lifespan study on rats showed reduced lifespan on rats fed fish oil [*90], possibly due to immune suppression by EPA. But rats are unlike humans in that they rarely die of cardiovascular disease and they have a lower capacity to synthesize the protective eicosanoid Prostacyclin (PCI3) from EPA. Other experiments, on mice, have shown extended lifespan with fish-oil. Fish oil even increased the life-extending benefits of calorie restriction on the mice [*91].
GLA supplements like evening primrose oil, or borage oil, might seem undesirable for normal people because of the danger that they would increase arachidonic acid. But studies have shown an increase in membrane DGLA and series 1 eicosanoids, with beneficial anti-inflammatory consequences [*92]. GLA can also lower blood pressure [*93].
To minimize lipid peroxidation, essential fatty acid supplements should be taken with no less than 500 mg of Vitamin E per day. Both gamma and alpha forms of tocopherol should be included [*94] for effectiveness. Vitamin E is an antioxidant which strongly prevents membrane peroxidation. The combination of Vitamin E and omega−3 fatty acid can reduce cancer risk while protecting heart cell membranes, brain cell membranes, immune-cell membranes and the receptor membranes that allow hormones to act. Vitamin E can protect essential fatty acids from lipid peroxidation in capsules and in the bloodstream, as well as in cell membranes. But for smokers, vitamin E may not be adequate to reduce oxidation of fish oil in LDL-cholesterol, and some medical authorities have cautioned against too much fish oil consumption by smokers [*95].
Ideally, essential fatty acids and Vitamin E should be formulated together in an air-tight capsule. Vitamin C helps maintain the antioxidant capabilities of Vitamin E. Boosting glutathione levels with N-acetyl-cysteine (NAC) and the use of other antioxidant nutrients can further protect against lipid peroxidation of essential fats. Glutathione not only regenerates both Vitamin E and Vitamin C that has been oxidized, it prevents formation of deadly hydroxyl free-radicals.
Optimum dietary benefit from fat for most people would come from a program of reduced total fat, reduced saturated and unessential fat, and increased proportions of omega−3 (relative to omega−6) essential fats. A high omega−3 oil like perilla oil might be a simple remedy for young people — and the best remedy for smokers. But as most people age, they will benefit most from CLA, GLA, and DHA supplementation combined with antioxidants (especially vitamin E) to protect these polyunsaturated essential fats from oxidation.
Fats are an important component of membranes in our hearts, brains, immune cells and most of the other tissues of our bodies. Since we need these fats, it is important to ensure that we have the right kind of fats, that we have enough of them and that we protect them with antioxidants.
![[GO TO BEN BEST'S HOME PAGE]](../homeback.gif) HOME PAGE
HOME PAGE